« Sapiens » dans « Homo sapiens » signifie « intelligent », « raisonnable », « sage », « sensé », et vient du latin sapere, qui veut dire « exhaler du goût, de la saveur », mais aussi « percevoir, sentir par le sens du goût », « avoir de l’intelligence, du jugement, des vues justes », « connaître », « comprendre », « savoir » (Gaffiot 1934). Homo sapiens, intelligent, doué de sensibilité et de sagesse, est animé de vie.
 [1]
[1]Figure 1 : Donax variabilis, une même espèce, de multiples apparences ou phénotypes : classer ou singulariser ? Source : « Coquina variation 3 » de Debivort, Wikimedia commons, CC-BY-SA.
Dans une conception matérialiste occidentale, tout être vivant est composé d’un ensemble d’atomes, organisés en molécules, elles-mêmes formant des cellules (des membranes délimitant un milieu intérieur en équilibre dynamique avec un milieu extérieur), le matériel nécessaire à sa réplication presque à l’identique se logeant au cœur de chacune d’elles. Dès sa naissance, l’homme est en interaction nécessaire, vitale, forcée, à la fois avec l’environnement physique dans lequel il puise matière et énergie, et avec son environnement social, c’est-à-dire les autres hommes et femmes en compagnie desquels il évolue et avec lesquels il entretient relation.Dans cette même conception matérialiste, ces interactions multiples sont sous-tendues par une vie chimique qui s’anime autour de son « génome » : un lot de chromosomes que lui ont transmis ses parents et qui se trouve (presque[1][2]) à l’identique dans chacune des 1014 cellules qui le composent. Ces 23 paires de chromosomes, d’origine maternelle pour moitié et paternelle pour l’autre, sont formées d’une longue molécule d’ADN (acide désoxyribonucléique) très compactée et elle-même composée d’une succession d’unités (« nucléotides ») appariées deux à deux et pouvant être de quatre sortes : A, C, G ou T. Ainsi, le génome humain contient trois milliards de paires de nucléotides dont seule une petite partie (1,5 % environ) est organisée en « gènes », des entités contenant chacune l’information nécessaire à la synthèse d’une « protéine » particulière. Pour l’essentiel, ce sont les protéines qui assurent l’activité vitale de nos cellules et donc le fonctionnement de notre organisme. Le génome humain contient quelque 20 000 gènes au total. Ce patrimoine génétique, matériel, chimique, nous le tenons de nos ancêtres, proches et lointains, chacun d’eux ayant transmis à ses descendants une moitié de son génome. De génération en génération, l’homme donc ne se « reproduit » pas, mais produit de la diversité, à travers un réassortiment génétique permanent, cœur du mode sexué de sa « reproduction ».
Il n’est pas rare d’entendre, de la part de généticiens, des propos tels que :
L’information contenue dans chacune de nos cellules raconte notre histoire individuelle, et aussi celle de tous les ancêtres dont nous descendons ; et nous transmettons cette information d’une génération à l’autre ; nous portons tous une archive de notre passé en nous, une archive généalogique. (Guérin 2011)
Les récits sur l’origine et l’évolution humaines ne sont cependant pas écrits dans notre séquence d’ADN; ce sont des études comparatives, des études centrées sur la différence entre les génomes des divers êtres vivants, qui les « révèlent » aux généticiens. Il s’agit dès lors de constructions scientifiques, humaines, de façons d’appréhender le monde, la nature et les autres hommes, dans l’illusion de (com)prendre, de « saisir », ce que nous sommes (et avons été), dans les dimensions transcendantes du temps et de l’espace. Cette illusion de se rapprocher au plus près d’une vérité sur la société, de la manière supposément la plus rationnelle et objective possible, est aussi celle qui sous-tend les histoires que la génétique nous révèle sur les rapports que nous entretenons avec nos ancêtres. Certes, notre ADN porte leur trace, mais la « lecture » de notre ADN peut-elle remplacer l’oralité des récits qui caractérise la plupart de nos histoires individuelles, ainsi que celle de l’humanité ?
L’historiographie humaine, entendue ici comme l’écriture de l’histoire humaine individuelle ou collective, produite par la génétique est particulièrement à l’œuvre ces dernières années pour les sociétés malgaches où, sortie de son contexte sociopolitique, elle touche une réalité sociale dont elle risque de mettre l’ordre en danger. Dans la présente analyse, à travers une approche latourienne, nous portons un regard critique et réflexif sur une production scientifique conduisant à une (re)définition scientifique de l’ancestralité, mise en circulation dans des sociétés malgaches où la construction identitaire individuelle et collective, et l’ordre sociopolitique qui en découle, sont sous-tendus depuis des siècles par une conception sociale et culturelle de l’ancestralité. L’intention de notre démarche interdisciplinaire est de juxtaposer un discours scientifique et toutefois narratif (Lyotard 1979) sur l’ancestralité, et la vision sociale, historique et vécue de celle-ci, ceci afin de donner à voir la mise en tension par une narration scientifique d’une conception locale de l’ancestralité. Nos préoccupations scientifiques se situent autour de la circulation des savoirs entre les différentes sphères de la société et la dynamique des imaginaires sociaux, entre imaginaires institués et instituants (Castoriadis 1996). L’objectif de ce texte n’est donc pas de « donner une leçon de réflexivité » à la communauté scientifique des généticiens qui ne semble pas en avoir besoin (Thoreau et Despret 2014), mais de susciter une mise en dialogue de la problématique de l’ancestralité entre des acteurs des sciences de la vie, d’une part, et d’autres membres de la société, d’autre part.
L’historiographie malgache revisitée par l’approche génétique.
Pourquoi cet intérêt des généticiens pour la population malgache ? La présence de l’homme à Madagascar, dont la connaissance n’est pas encore stabilisée, apparaît récente : elle ne daterait que de 4000 ans environ[2][3]. Cette « jeunesse » du peuplement de Madagascar est d’autant plus étrange qu’Homo sapiens, né il y a 200 000 ans en Afrique, aurait commencé sa migration hors du continent africain, par la corne de l’Afrique, 70 000 ans BP environ[3][4]. Aussi le peuplement de l’île reste-t-il une « énigme » pour les scientifiques issus de disciplines diverses (principalement la linguistique, l’archéologie et la génétique) qui cherchent à en donner une date plus précise, à en déterminer l’origine, la nature et le contexte historique.
La curiosité associée à cette question provient aussi, en grande partie, du contraste entre l’homogénéité linguistique et la grande diversité des types somatologiques rencontrés. La langue malgache, parlée dans l’ensemble de l’île, appartient à une famille de langues austronésiennes d’Asie du Sud-Est : la famille des langues barito, aujourd’hui parlées au sud de l’île de Bornéo (Dahl 1951). Otto Dahl (1991) soutient que la thalassocratie malaise de Srivijaya, au sud de Sumatra, est à l’origine de la migration proto-malgache vers le 7e siècle. Par ailleurs, la langue malgache contient un nombre considérable de termes d’origine bantoue dont la source exacte en Afrique n’a pu être identifiée jusqu’à présent. Ainsi, la migration proto-malgache d’origine austronésienne s’inscrit dans une longue histoire de contacts entre l’Afrique et l’Asie du Sud-Est. Dans le cadre de l’hypothèse de la double ascendance afro-asiatique, la question du métissage entre les populations proto-malgaches bantoue et austronésienne reste toujours d’actualité. Alors qu’Alfred Grandidier attribue un rôle annexe et tardif à l’Afrique dans le peuplement de Madagascar, Gabriel Ferrand fait précéder l’apport austronésien par le monde africain bantou (Grandidier 1901, Ferrand 1909). Plus récemment, Alexander Adelaar (2012) a privilégié une hypothèse selon laquelle les premiers colonisateurs de Madagascar constituaient déjà une population métisse austronésienne-bantoue. Cela expliquerait l’homogénéité linguistique à Madagascar et le fait que, dans la langue malgache, les termes d’origine bantoue désignant les plantes et les animaux domestiques de base sont plus importants et leur signification bien plus fidèlement conservée que les termes d’origine barito[4][5].
Plusieurs sources écrites datant du début de notre ère mentionnent la côte orientale africaine. Dans le Périple de la Mer Érythrée, récit gréco-romain d’exploration maritime datant du 1er siècle après Jésus-Christ, Madagascar apparaîtrait sous le nom de « Menuthias ». Les Austronésiens, au moyen de radeaux ou de canoës à balancier et faisant usage de contre-courants équatoriaux saisonniers, auraient apporté des épices en échange d’objets manufacturés gréco-romains (poterie, perles de verre…) (Blench 2007, 2010). En se basant sur des découvertes archéologiques et sur divers récits de voyage de géographes arabes datant du 10e au 12e siècle, notamment le Livre des Merveilles de l’Inde[5][6], Claude Allibert (2005) fait l’hypothèse d’une traite d’esclaves africains par les Austronésiens au 10e siècle, en direction des îles comoro-malgaches : des habitants de la côte orientale africaine étaient emmenés par des Austronésiens, non pas sur leurs propres territoires en Asie, mais pour être exploités quasiment sur place, au voisinage de Kanbalu (région de Zanzibar), suffisamment à l’écart des côtes africaines pour qu’ils soient contrôlables. Dans une vision englobante, Philippe Beaujard (2012) parle de l’ensemble africain touché par le tropisme indianocéanique, dans une vision de l’Afrique comme une périphérie de l’océan Indien et des villes swahilies comme des provinces, depuis le 8e siècle.
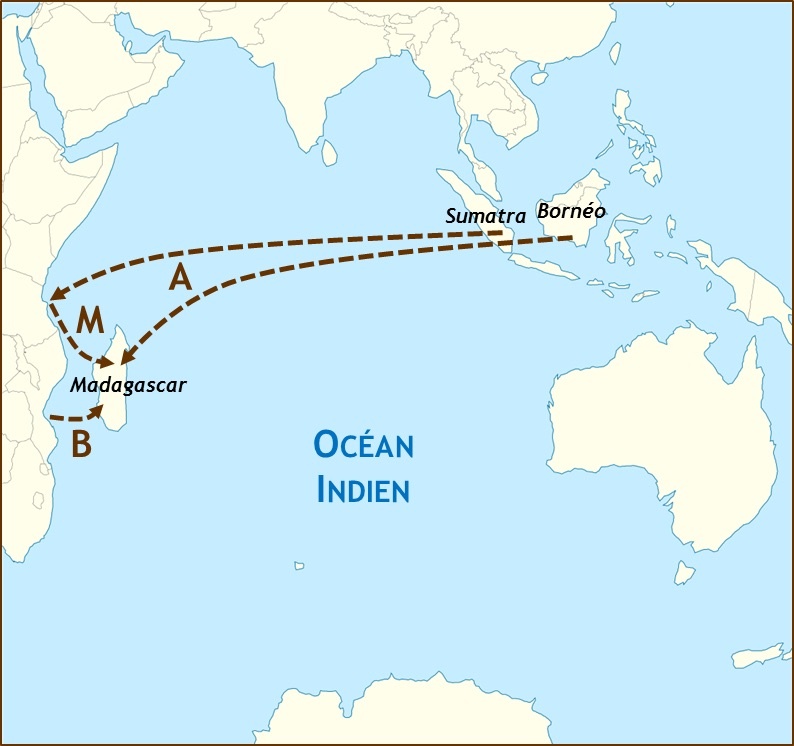 [7]
[7]Figure 2 : Schéma de différentes hypothèses sur l’origine du peuplement de Madagascar. La migration proto-malgache est-elle d’origine austronésienne (A), bantoue (B), ou l’île a-t-elle initialement été peuplée par une population métisse austronésienne-bantoue qui se serait préalablement formée sur la côte orientale africaine (M) ? Figure adaptée de « Indian Ocean laea location map » de Tentotwo, Wikimedia commons, CC-BY-SA).
Parmi les différentes manières de déployer un argumentaire pour produire un récit du peuplement de l’île, l’approche génétique s’intéresse à l’étude du polymorphisme génétique malgache, qu’elle recherche dans l’ADN mitochondrial[6][8] (d’origine maternelle), le chromosome Y (d’origine paternelle) ou l’ADN autosomal (tous les chromosomes à l’exception des chromosomes sexuels X et Y). Depuis 2005, de nombreux généticiens se sont penchés sur la question des origines du peuplement de la Grande Île, en récoltant des échantillons de plus en plus nombreux, évoluant avec les techniques de traitement et d’analyse de plus en plus sophistiqués et performants, notamment du point de vue statistique (Hurles 2005, Tofanelli 2009, Razafindrazaka 2010, Cox 2012, Poetsch 2013, Pierron 2014). Ces travaux se concentrent essentiellement sur une analyse à trois niveaux : l’ascendance austronésienne ou africaine, l’hérédité maternelle ou paternelle de cet héritage, et le caractère « ethnique » de la diversité génétique. Ces analyses, que nous ne détaillons pas ici, sont menées au sein de divers groupes dits « ethniques », eux-mêmes distingués comme « gens des Hautes Terres » ou « côtiers »[7][9]. Les recherches s’accordent à peu près toutes sur un brassage, une hybridation génétique relativement récente (un millier d’années environ) entre les populations austronésienne et bantoue : leurs contributions génétiques relatives sont à peu près égales, avec toutefois de petites prépondérances ici et là qui offrent des opportunités d’interprétation quant aux processus migratoires. Pour ceux qui penchent pour une origine asiatique, il s’agit d’un petit nombre de colons initiaux arrivés en un seul processus migratoire (Razafindrazaka 2010), ou d’une colonisation datant de 1200 ans par une trentaine de femmes d’origine indonésienne à 93 %, avec en outre l’hypothèse d’un seul bateau qui aurait involontairement fait naufrage à Madagascar (Cox 2012)[8][10]. Les auteurs soulignent que « la contribution indonésienne (à travers la langue, la culture et les gènes) continue de dominer la nation de Madagascar aujourd’hui » (ibid., p. 2767). Pour d’autres chercheurs, si l’analyse de l’ADN mitochondrial montre un héritage génétique d’origine asiatique plus important, c’est l’ascendance africaine qui domine largement dans les analyses du chromosome Y et de l’ADN autosomal, « ce qui n’est pas surprenant compte tenu de l’apparence africaine des Malgaches » (Poetsch 2013, p. 1427).
Aussi, plus récemment, et de plus en plus, les généticiens recherchent dans les génomes d’une population, les traces d’une adaptation à un environnement, à un mode de vie particulier, les traces d’une sélection naturelle. C’est le cas de l’étude conduite par des généticiens sur l’origine des chasseurs-cueilleurs Mikea à Madagascar (Pierron 2014). Ces derniers sont considérés par certains comme une survivance du peuplement pré-bantou de l’île. La présence de chasseurs-cueilleurs dans le sud de l’île est rapportée dans les récits de voyageurs européens jusqu’au 16e siècle, et confirmée par des données archéologiques. Les raisons de leur disparition après le 16e siècle sont inconnues. Les Mikea sont aujourd’hui l’objet d’une controverse : sont-ils les descendants de chasseurs-cueilleurs anciens qui auraient maintenu leur mode de vie à travers le temps, ou d’agriculteurs sédentaires qui auraient été « renvoyés » vers la forêt pour des raisons politiques et économiques (sous la pression de la royauté Sakalava ou de la colonisation française) ? Des travaux récents montrent que les Mikea présentent le même profil de diversité génétique que leurs voisins pêcheurs semi-nomades (les Vezo) et que les éleveurs et agriculteurs Antaimoro, ce qui signale une hybridation récente entre Bantous et Austronésiens, la part bantoue représentant plus de 60 % du fond génétique et la part austronésienne environ 30 %. Les Mikea seraient donc les descendants d’une population d’agriculteurs qui serait retournée vers la forêt. Mais les auteurs de cette analyse ne se sont pas arrêtés là. Ils ont recherché des signatures génétiques particulières, caractéristiques d’une sélection naturelle récente, attribuable à une adaptation au mode de vie chasseur-cueilleur. Chez les Mikea, cette recherche pointe des gènes en relation avec le fonctionnement neuronal, l’immunité adaptative, la contraction des muscles striés et le métabolisme d’aliments particuliers, des résultats qui pourraient être en lien avec le mode de vie chasseur-cueilleur ; ces résultats préliminaires nécessitent des recherches complémentaires pour caractériser les effets phénotypiques (visibles, mesurables) de ces variants génétiques. Quoi qu’il en soit, la recherche, chez les Mikea, d’une signature génétique d’adaptation à l’environnement aboutit à une différenciation génétique statistique de la population Mikea par rapport aux populations voisines Vezo et aux Antaimoro. Étant une forme de discrétisation sociale[9][11], ces travaux peuvent bien entendu être porteurs de perspectives de divers ordres (foncier, identitaire, etc.).
Une telle discrétisation des sociétés malgaches par la génétique contemporaine inquiète et suscite de nombreuses questions : comment est-il possible que l’on cherche à connecter des gènes à des caractéristiques d’ordre social ou culturel ? Comment en est-on arrivé là ? De quoi parle-t-on exactement ? Que représente la diversité génétique humaine ? Où se loge-t-elle ? Quelles conséquences ces recherches vont-elles avoir sur l’ordre social à Madagascar ? En d’autres termes, comment et dans quelle mesure peut-on relier une apparence physique, voire un comportement, à un patrimoine génétique ? Et pourquoi choisit-on de le faire auprès de populations telles que les sociétés malgaches, dont on sait combien elles sont hiérarchisées et à quel point l’ordre social repose sur une ancestralité certes basée sur la filiation, mais également sur le territoire et l’alliance ? Enfin, comment l’ADN peut-il avoir une telle valeur narrative, et quelles conséquences cela peut-il avoir sur une historiographie individuelle et collective, sous-tendue jusqu’ici par une tout autre conception de l’ancestralité ? Pour apporter des éléments de réponse à un tel questionnement, un détour par les recherches actuellement menées sur le génome humain s’impose : l’apparence physique et le comportement humain sont précisément au cœur des grands projets scientifiques contemporains dits « génomiques » (qui ont pour objet d’étude le génome humain).
La relation entre ce que les yeux croient voir et le patrimoine génétique.
La diversité génétique humaine est étudiée à travers le monde dans le cadre de grands consortiums scientifiques internationaux et d’entreprises privées, tels que Human Genome Project HGP[12], HUman Genome Organization HUGO[13], HapMap[14], 1000 Genomes[15], ou encore Human Genome Diversity Project[16] et Genographic Project[17]. Ceux-ci affichent comme objectif principal l’amélioration de la santé en mappant, localisant et identifiant les variants génétiques impliqués dans les maladies dites « complexes » (d’origine multigénique), comme l’hypertension, l’asthme, le cancer ou la démence. Dans le cadre des Genome-Wide Association Studies (GWAS), on recherche la corrélation entre la prévalence d’une maladie, ou d’un trait complexe, et une caractéristique, un marqueur génétique, par exemple une mutation. Les grands projets comme HapMap tentent de dresser un catalogue des variations génétiques les plus fréquentes chez l’homme. Ils ont pour finalité de fournir aux chercheurs les informations qui leur permettront de relier ces variations génétiques aux risques de contracter telle ou telle maladie. À terme, c’est une médecine personnalisée qui est visée, au niveau de la prévention, du diagnostic et du traitement (domaine de la pharmacogénomique)[10][18].
Du point de vue génétique, chaque personne est caractérisable par son génotype (sa composition génétique, sa séquence d’ADN) et par son phénotype (ses caractères observables : morphologiques (externes), anatomiques (internes), physiologiques (en rapport avec son fonctionnement physique et biochimique) ou moléculaires). L’homme a probablement toujours relié génotype et phénotype sans le savoir. Ainsi, nous disons : « tel père, tel fils », « les chiens ne font pas des chats », ou encore, « the apple doesn’t fall far from the tree ». Gregor Mendel a proposé les lois de la génétique qui définissent la manière dont le patrimoine génétique se transmet de génération en génération en s’intéressant notamment à la couleur des fleurs de pois, à la forme des gousses, etc. Cependant, le phénotype n’est que partiellement déterminé par le génotype. Le phénotype — l’apparence — est aussi influencé par l’environnement (le régime alimentaire, par exemple). De plus, deux séquences identiques d’ADN, deux successions identiques de nucléotides A, C, G ou T, peuvent aboutir à des expressions différentes en protéines[11][19]. La relation entre génotype et phénotype est donc extrêmement complexe. Les projets sur le génome humain sont cependant fondés sur « la croyance selon laquelle tout ce que nous voulons savoir sur les êtres humains est contenu dans la séquence de leur ADN » (Lewontin 1992, p. 9). Or, nous savons que la maladie, et la souffrance humaine en général, est davantage le résultat de la pauvreté (et son cortège de malnutrition, surcharge de travail, accès limité aux soins, etc.) que d’origine génétique. Les gènes ne déterminent pas l’hérédité : ils y participent avec l’environnement (Jablonka 2005). Le danger d’une telle croyance dans le déchiffrage du « grand livre de la vie » est de renforcer l’idéologie centrée sur le déterminisme biologique comme explication des différences sociales et individuelles. Pourtant, chaque homme est unique, singulier ; les hommes sont tous différents, mais aussi égaux en droits dans l’idéal proclamé par la Déclaration des droits de l’Homme.
La singularité humaine traitée par l’approche comparatiste de la génétique des populations.
Si les grands projets de génomique humaine relèvent de la génétique humaine, ils relèvent aussi et surtout de la génétique des populations, puisqu’ils sont avant tout fondés sur une approche comparatiste des groupes humains. La génétique des populations étudie l’évolution du patrimoine génétique au cours des générations. Elle a pour base théorique la génétique mendélienne (l’hérédité des caractères biologiques), la théorie de l’évolution de Darwin (« survival of the fittest », ou « survie du plus adapté ») et la théorie neutraliste de l’évolution de Kimura (« survival of the luckiest », « survie du plus chanceux ») trop souvent oubliée. Cette dernière prend pourtant en compte la neutralité de la plupart des mutations et la dérive génétique, causée par des phénomènes aléatoires, comme le hasard des rencontres entre un spermatozoïde et un ovule particuliers. Ainsi, la diversité génétique dans une population n’est pas forcément représentative de la génération antérieure, cet effet étant d’autant plus important que la population est petite.
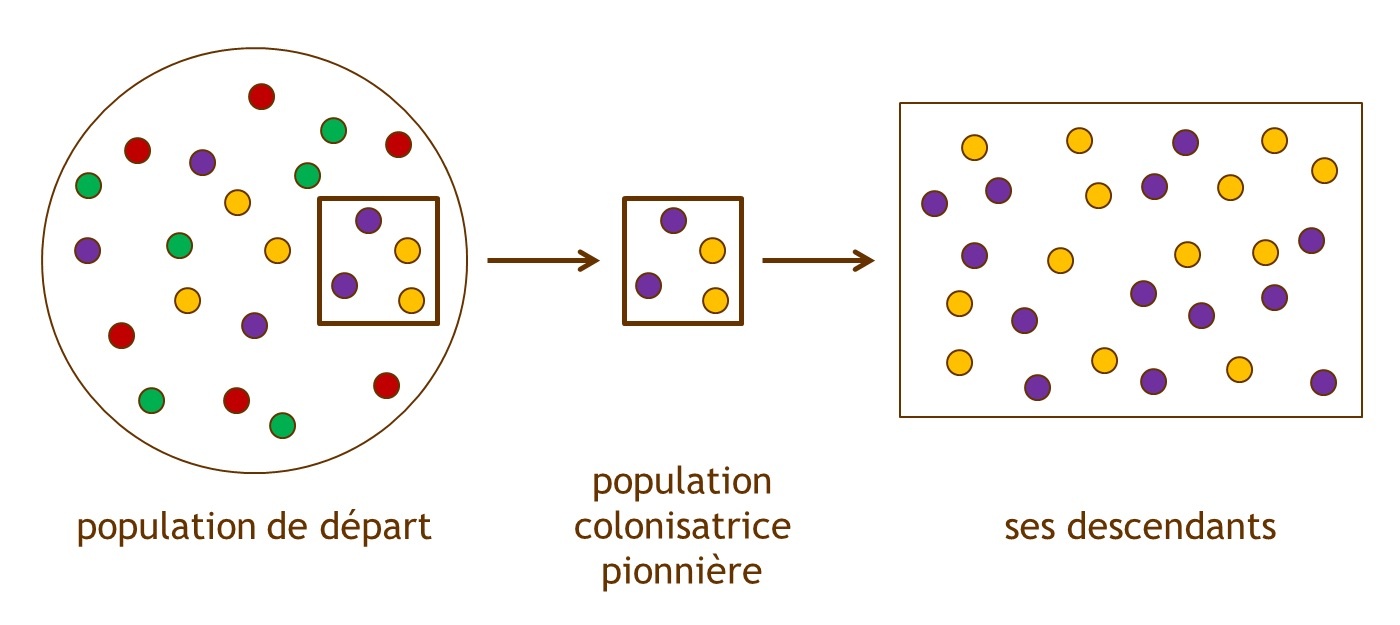 [20]
[20]Figure 3 : L’« effet fondateur », exemple d’une dérive génétique. Les ronds colorés représentent les différents variants génétiques d’un même gène (allèles). Lorsqu’une partie de la population émigre pour coloniser un nouveau territoire, la diversité génétique est diminuée dans la population issue de cette émigration et ne représente donc pas celle de la population initiale. La dérive génétique produit, au hasard, une variation de la fréquence des différents allèles dans la population des descendants, et ce en absence de toute sélection naturelle. Source : Sonia Dheur.
Les hommes sont tous différents, uniques, mais dans quelle mesure, d’un point de vue génétique ? Les génomes de deux personnes prises au hasard dans la population mondiale sont identiques à 99,9 %. Autrement dit, ils diffèrent d’un nucléotide sur 1000 en moyenne (la diversité génétique humaine trouve principalement son origine dans les erreurs de copie qui se glissent au moment de la réplication de notre ADN avant chaque division cellulaire). Il existe différents types de mutations, la substitution (le remplacement d’un nucléotide par un autre) étant la mutation la plus fréquente, et donc le type de variation génétique le plus fréquent entre deux individus. On l’appelle « polymorphisme nucléotidique » ou SNP (single-nucleotide polymorphism). Parmi les 10 millions de SNP dans le génome humain, quelques-uns seulement seraient liés à des maladies, la plupart du polymorphisme étant neutre (sans incidence sur les protéines exprimées et ne correspondant donc à aucune différence observable). Aujourd’hui, on dispose de « puces à ADN », une technologie qui permet de déterminer plus de 700 000 SNP à partir d’un échantillon d’ADN. On est donc capable de « génotyper » chaque individu, de le caractériser génétiquement.
Pour investiguer la diversité génétique humaine, les grands projets génomiques cités plus haut s’adressent à des hommes et des femmes répartis dans le monde. À partir d’un prélèvement de salive contenant quelques cellules buccales, des machines (contrôlées par des ordinateurs) en extraient l’ADN, l’amplifient, le fragmentent et l’hybrident sur des puces à ADN, en prélèvent les signaux d’hybridation et en déterminent les variants pour les 700 000 SNP. Ces données sont ensuite traitées par de puissants programmes statistiques, calculant les distances génétiques entre les personnes analysées, dans un espace à n dimensions. Ces programmes cherchent ensuite, dans cet espace, les directions correspondant aux variations les plus importantes (les « composantes principales »), afin de projeter les résultats dans un espace à deux dimensions, appréhendable par nos sens. Les personnes sont alors représentées par des points dont la répartition graphique en nuages colle extraordinairement à la répartition géographique des individus dans l’espace, et ce avec une très grande résolution (Novembre 2008). Ainsi, l’analyse ADN en composantes principales d’un millier d’individus issus de 14 comtés de l’Estonie a révélé une distribution graphique de la diversité génétique similaire à la répartition géographique, spatiale, de ces 14 populations (Nelis 2009). Pourquoi ? « Cupid has short wings », répond Luigi Luca Cavalli-Sforza, généticien des populations et inventeur de la géographie génétique (Manni 2010). Autrement dit, nos partenaires sexuels sont en majorité proches de nous dans l’espace, l’endogamie de localité faisant ainsi de nos voisins nos parents.
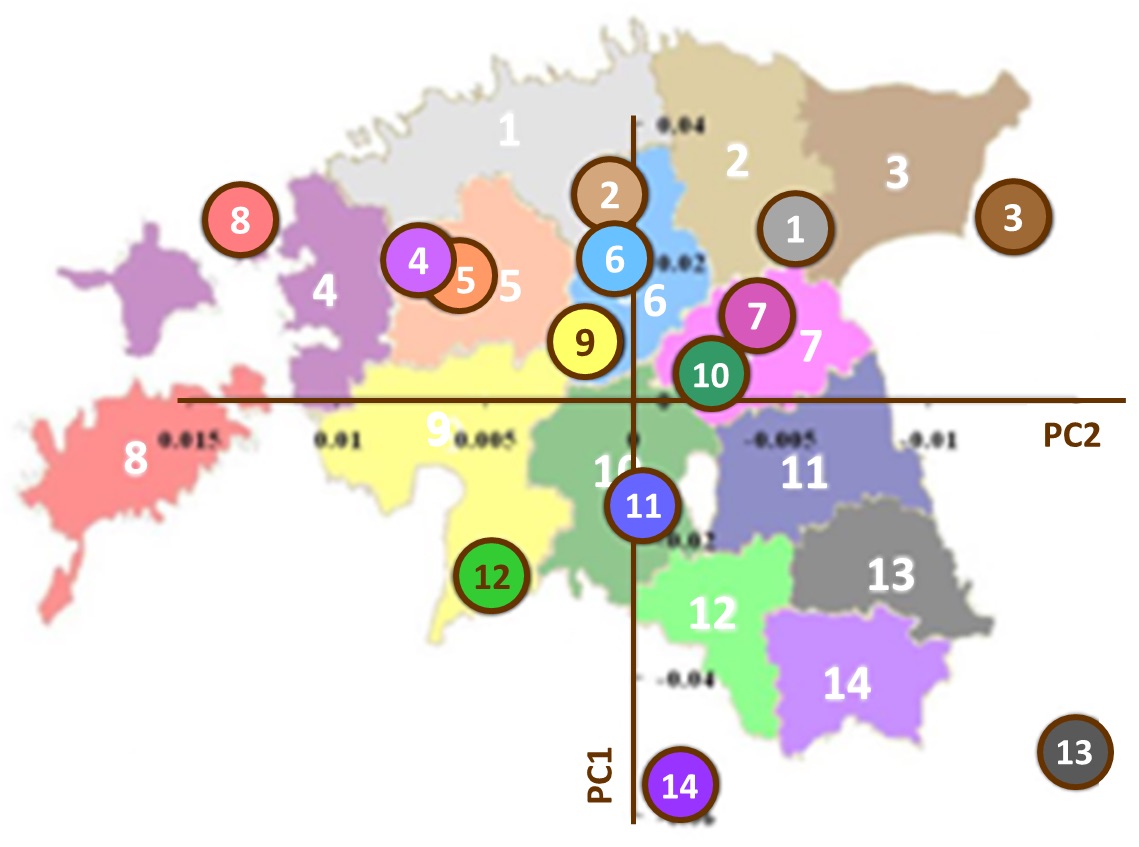 [21]
[21]Figure 4 : Structure génétique de l’Estonie. Analyse en composantes principales (PC1 et PC2) des données génétiques issues d’un millier de personnes natives de 14 comtés d’Estonie. Les cercles colorés et numérotés représentent chacun la médiane des valeurs pour tous les individus d’un comté donné pour les deux composantes principales. La superposition de ces données génétiques avec une carte géographique de l’Estonie montre combien les proximités génétique et géographique sont corrélées. Figure adaptée de Nelis et al. 2009, p. e5472.
En génétique des populations, la population est définie comme un groupe d’individus habitant un même territoire et présentant une communauté de reproduction, une relative endogamie. Relativement à la diversité génétique humaine globale (portant sur 0,1 % du génome), les différences génétiques entre populations ne représentent que 15 % de celle-ci (7 % en moyenne entre deux populations) ; 85 % de la totalité de la diversité génétique humaine se trouve donc au sein de chaque population. Autrement dit, la diversité génétique humaine est principalement due à des différences individuelles (Lewontin 1972, Barbujani 1997). Entre populations, la diversité génétique se traduit par de petites différences dans la fréquence de certains SNP par rapport à une autre population, aucun SNP n’étant exclusif d’une seule population. Il n’existe donc aucun gène pour lequel une population (géographiquement définie) contiendrait un variant génétique avec une fréquence de 100 % et pour lequel une autre population aurait un autre variant dans 100 % de ses membres (Feldman et Lewontin 2008). Ces SNP, plus ou moins fréquents dans certaines populations, sont essentiellement stochastiques, dus à la dérive génétique, une évolution causée par des phénomènes aléatoires, et très minoritairement le fruit d’une adaptation, voire d’un perfectionnement, dus à la sélection naturelle. Ils sont neutres pour leur grande majorité ; ils ne se situent pas dans une portion de l’ADN contenant un gène, n’ont aucune incidence sur les protéines exprimées, et n’ont donc un effet ni sur l’apparence ni sur l’aptitude. Dans de très rares cas seulement, ces SNP sont localisés dans des gènes et peuvent influencer le métabolisme ou l’apparence physique[12][22].
Rappelons que nous sommes tous « d’origine africaine ». D’après l’hypothèse « out-of-Africa », Homo sapiens est né en Afrique il y a environ 200 000 ans. Il a ensuite progressivement peuplé les différents continents. Par un effet appelé « effet fondateur », l’importante diversité génétique africaine s’est progressivement diluée au cours des migrations successives, un petit groupe se séparant chaque fois d’un ensemble plus grand, donc plus diversifié, à chaque colonisation d’un nouveau territoire. Cependant, l’origine commune et relativement récente de l’homme moderne explique notre homogénéité génétique (notre identité à 99,9%)[13][23]. Notre origine commune et récente explique aussi le fait que la plus grande part du polymorphisme génétique total se trouve au sein de chaque population. Mais c’est en traquant les petites différences entre des individus issus des populations du monde entier que les chercheurs en sciences de la vie (re)tracent « l’aventure humaine », une aventure à travers le temps et l’espace qui les concernent et les intéressent. Et pendant ce temps, les chercheurs en sciences humaines et sociales s’inquiètent de la réémergence du concept de « race ».
Lorsque les Européens, dès le 15e siècle, réduisaient en esclavage les populations africaines qu’ils considéraient comme « inférieures », ils sanctionnaient l’apparence des fondateurs de l’humanité tout entière. La notion moderne de « race » apparaît tard dans l’histoire des hommes. Elle se renforce au 18e siècle, où elle est en lien avec le grand processus de colonisation, et sert à justifier idéologiquement l’immense violence déployée pour coloniser le monde et en drainer les richesses vers le continent européen. Jusqu’au milieu du 20e siècle, les théories racistes vont s’appuyer sur des arguments biologiques : les races existent parce que les hommes sont fondamentalement différents d’un point de vue physique, anatomique et génétique. Le concept de « race » est une construction sociopolitique. La science montre que le terme de « race » comme entité génétique indépendante n’existe pas, ce qui ne veut pas dire que nous sommes tous pareils (Jordan 2010).
Pour les généticiens des populations, depuis la théorie synthétique de l’évolution datant des années 1930-40, ce sont les relations entre les individus (la généalogie, l’endogamie, la compétition), et non des propriétés individuelles intrinsèques, qui déterminent l’appartenance à un groupe. En aucun cas, il n’est attendu que tous les membres du groupe partagent une caractéristique, ni génétique ni phénotypique, propre à ce groupe (Gannett 2014). Les différences génétiques entre populations sont cependant avérées en termes de fréquences de certains variants génétiques : nous sommes capables de discrétiser du point de vue génétique les populations habitant l’Europe, l’Afrique ou l’Asie (Shriver 2004). Bien qu’il aille de soi que ces différences ne peuvent sous-tendre aucune hiérarchisation, ne faut-il pas toutefois dissocier le discours scientifique et le discours moral ?
Le choix de voir en d’autres humains nos égaux, dont la seule existence nous impose l’obligation de respect, de bienveillance et d’assistance, est un commandement de la conscience, qui n’a pas besoin d’un aval de la science. La science peut nous instruire sur ce qu’est le monde ; elle est incapable de nous dire comment nous devrions nous y conduire. (Stoczkowski 2006, p. 48)
Dans sa constitution de 1950, au lendemain de la Seconde Guerre mondiale, devant la révélation des crimes nazis, l’UNESCO affirmait l’unité fondamentale de l’espèce humaine. Dans le même temps, on investissait la science d’une mission : pénétrer les profondeurs du patrimoine génétique (génotype) pour démentir ce que les yeux croient voir, l’apparence, le phénotype. On attendait de la science qu’elle mette fin à l’idée que les différences de culture sont sous-tendues par des différences de nature. Mais la période où les généticiens se sont concentrés sur l’unité génétique humaine a été très courte. Les travaux sur le génome humain se sont très vite et de plus en plus orientés vers l’exploration de la diversité génétique au sein de notre espèce. Si bien que nous sommes capables de distinguer génétiquement les individus ayant des ancêtres issus d’Europe, d’Afrique ou d’Asie (Stoczkowski 2006).
Le façonnement du temps et de l’espace par la génétique contemporaine.
Les généticiens définissent quatre « groupes géographiques ancestraux » (biogeographical ancestries ou BGA), c’est-à-dire quatre groupes majeurs d’ascendance, « fondateurs » : les « Européens », les « Amérindiens », les « Africains sub-sahariens » et les « Asiatiques orientaux ». Ces BGA représenteraient les groupes fondateurs des populations actuelles. Et c’est là où le bât blesse… Quels groupes fondateurs, quels groupes ancestraux ? Car en réalité, la divergence génétique s’étend dans l’espace (comme dans le temps) de façon clinale : lente, progressive, et non discrète. Les fréquences des SNP varient ainsi de façon continue à travers une aire géographique (Fujimura 2014). Lisa Gannett (2014), historienne et philosophe des sciences, l’explicite très simplement : par exemple, un échantillonnage à la périphérie du continent eurasiatique (en Irlande à l’ouest et au Japon à l’est) va donner des groupes plus distincts génétiquement que si les échantillons étaient prélevés dans deux populations plus centrales de la même aire, mais en aucun cas cela ne signifie que les groupes de la périphérie sont des descendants purs de lignages primordiaux, et que les groupes plus centraux sont des hybrides de ces lignages ancestraux. Pour Gannett (2014), ces catégories sont des catégories locales qui correspondent aux régions du monde où beaucoup d’Américains cherchent la trace de leurs ancêtres. Il ne s’agit pas d’une catégorie correspondant à une classification biologique ou anthropologique. Le concept d’« ancestralité continentale » reste une catégorisation raciologique, car elle est fondée sur l’idée de pureté d’un groupe ancestral ; c’est une construction édifiée sur le concept de « race », construite socialement et politiquement dans la tradition scientifique et philosophique européenne.
Ces catégories biogéographiques sont utilisées dans la recherche biologique et biomédicale comme alternative aux catégories raciales et ethniques. Rappelons qu’aux États-Unis, les concepts de « race » et d’« ethnie » sont des catégories officiellement reconnues. Lors du recensement de la population, chacun s’auto-déclare Blanc, Noir, Amérindien… Et les scientifiques dont les recherches sont soutenues financièrement par le NIH (National Institute of Health) se doivent d’utiliser ces mêmes catégories afin de s’assurer que les « minorités » sont incluses dans les essais cliniques. Pour Gannett (2014), le concept d’« ancestralité continentale » n’est donc qu’un repackaging (reconditionnement) de la notion de « race » : il permet d’éviter le mot « race » au passif, trop chargé politiquement. Mais les choses sont plus compliquées encore, car l’appartenance à ces nouvelles catégories d’ancestralité géographique est aujourd’hui revendiquée comme principe d’identité individuelle et collective. Ceux que l’on classait dans les « minorités ethniques » (les Afro-américains, les Amérindiens…) affirment de plus en plus leur identité avec fierté, grâce à des entreprises qui proposent de reconstituer les lignées ancestrales par l’analyse génétique[14][24]. L’identité, en tant que représentation socialement construite, réclame des certitudes scientifiques[15][25]. Mais du point de vue biologique, même si les fréquences des gènes et les traits phénotypiques varient tous deux de façon clinale à travers l’espace géographique, les caractéristiques génétiques et phénotypiques ne varient pas de façon concordante, différents traits présentant différents schémas de variation dans l’espace. De plus, un ensemble de caractères biologiques définis par un code racial (tels qu’une couleur de peau, une texture de cheveu, des traits faciaux) ne co-varient pas ; ils varient de façon indépendante à travers l’espace.
En raison de ces variations clinales non concordantes, notre espèce n’est pas subdivisée en groupes raciaux ou continentaux discrets, distincts génétiquement et biologiquement homogènes. Les populations humaines tendent plutôt vers une similarité génétique par rapport aux populations voisines, indépendamment des frontières continentales ; le degré de similarité génétique étant inversement corrélé à la distance géographique. (Fujimura 2014, p. 210)
Dans le débat qui oppose les conceptions constructiviste et déterministe de la notion de « race », notre positionnement est constructiviste. Nous partageons l’idée suivant laquelle la notion de « race » ou d’« ethnie » est le résultat d’une fabrication socio-historique dynamique, une idée par ailleurs largement admise dans la communauté des généticiens. À l’instar de Fujimura et al. (2014), nous considérons que les chercheurs en sociologie et en histoire sont arrivés à un consensus selon lequel les catégories de « race » ou d’« ethnie » varient dans le temps et dans l’espace et se construisent en dialogue étroit avec des processus politiques, sociaux, culturels et psychologiques. Par ailleurs, nous sommes convaincus que, malgré la complexité génétique à laquelle ils sont confrontés, les chercheurs seront capables d’assigner un fond génétique (ou épigénétique) particulier à beaucoup de nos traits physiques, voire nos comportements, avec le temps. Cette perspective n’est cependant pas à confondre avec une vision déterministe du concept de « race » car, comme Hartigan (2008), nous sommes certains que la génétique ne sera jamais capable ni d’affirmer ni d’infirmer le fondement social et culturel de la race. La raison en est simple : les codes de la race (traits physiques, comportements, compétences, aptitudes, etc.) sont des définitions sociales et culturelles[16][26]. Les défenseurs de la vision constructiviste de la race cherchent trop souvent à s’appuyer sur les sciences dites dures pour démentir l’existence d’une conception déterministe de la race. Dans nos sociétés occidentales, où nous avons la longue tradition culturelle de diviser nature et société, la biologie dont fait partie la génétique est une science, donc une production sociale et culturelle, et non le reflet d’une réalité naturelle, la nature en soi. Science et société n’ont pas à être opposées : elles sont toutes deux une production humaine. Les conceptions constructiviste et déterministe ne s’opposent pas réellement, parce que les généticiens trouveront probablement toujours un fondement génétique (ou épigénétique) aux codes qui définissent la race ou l’ethnicité. Le fait que des codes raciaux tels que la couleur de la peau aient un fondement génétique ou pas nous éloigne de questions beaucoup plus prégnantes sur les raisons qui font que ces traits particuliers continuent de diviser la société en catégories hiérarchisées, conduisant à des injustices et des inégalités socioéconomiques, politiques, etc.
Les travaux mentionnés, ainsi que l’exposé des techniques, mais aussi des perspectives de la génétique contemporaine — en particulier celles offertes par la génomique —, en montrent la puissance de l’approche dans le champ d’études de l’histoire des groupes humains. Ils montrent aussi que, si les résultats concernant des ascendances d’individus peuvent générer une généalogie génétiquement traçable, une telle production n’en obéit pas moins à une construction mentale. La jeunesse du brassage génétique observé à Madagascar confère à sa population un caractère exceptionnel et précieux aux yeux des généticiens. Les caractéristiques génétiques particulières des génomes malgaches récemment mixés en font des outils de développement de l’analyse génomique en général, applicables à l’ensemble de la population humaine. D’une part, la population malgache est l’exemple de choix pour les recherches sur la détermination de l’origine des populations, comme l’explicite très bien le titre de Poetsch (2013) : « Determination of population origin : a comparison of autosomal SNP, Y-chromosomal and mtDNA haplogroups using a Malagasy population as example ». D’autre part, l’analyse génétique de populations malgaches telles que les Mikea permet de faire progresser plus efficacement les recherches sur les mécanismes généraux de la sélection naturelle. Mais, comme nous l’avons vu, la méthode utilisée dans ces recherches est toujours comparatiste. Elle implique des opérations de regroupement des personnes dont on étudie le génome, sous des étiquettes diverses : sur une base continentale (bantou/austronésien), ethnique (Mikea, Vezo, etc.) ou autre (gens des Hautes Terres/côtiers). La nomination des populations dans les analyses génétiques n’est pas neutre ; elle touche la question sensible de l’identité, particulièrement ancrée dans l’ancestralité à Madagascar : une conception complexe qui va bien au-delà de la simple parenté génétique et qui structure l’ensemble de la société. Aussi faut-il considérer le contexte sociopolitique dans lequel s’inscrivent la conception de telles études et la diffusion des résultats scientifiques de cette approche qui veut enrichir l’historiographie malgache par la génétique.
La question des origines et la réalité sociale à Madagascar.
La construction identitaire malgache est spécifique, dans le sens où elle donne priorité à la question des origines : on cherche à situer son interlocuteur dans la hiérarchie sociale pour voir dans quelle mesure il est susceptible d’appartenir à son propre réseau de parenté. Traitant du champ de l’ancestralité, Paul Ottino (1998) distingue trois modes de parenté : la parenté identitaire d’ancestralité, la parenté par le patrimoine indissociable de la première, et la parenté de parentèle produite par l’affinité et l’alliance. La parenté d’ancestralité est définie par rapport à un ancêtre regardé comme l’ancêtre d’origine ; elle est à la fois une parenté de descendance et de localité. Idéalement, Ego réside sur la terre ancestrale, terre défrichée par l’ancêtre et où se trouve son tombeau. Cet aspect de l’ancêtre d’origine, à la fois progéniteur et fondateur (dont on tient les droits), explique que la parenté d’ancestralité soit indissociable de la résidence, et donc de la parenté par le patrimoine ou parenté patrimoniale. Partout à Madagascar, l’idée de parenté est indissociable de celle de propriété, d’une propriété familiale ou ancestrale transmise héréditairement, dont les héritiers ne sont que les gardiens et dont le premier devoir est de la perpétuer en la transmettant à leur tour à leurs descendants. Les modes de parenté ancestrale et patrimoniale sont à la base de l’identité principale, à la fois rituelle et sociale d’Ego. Ils décident de l’affiliation, ou de la non-affiliation, à des catégories ou groupements d’ancestralité, ou à des groupes de co-héritiers. Les enjeux que ces modes de parenté constituent sont cruciaux et expliquent l’intensité des émotions qui accompagnent les situations qui les mobilisent. À la différence des deux premières, la parenté de parentèle, indissociable du mariage, peut être considérée comme relativement ouverte, à l’exception des sociétés malgaches favorisant l’endogamie. Elle a pour vocation de mettre en relation de proche en proche des groupements de descendance, d’ancestralité, ou des groupes de cohéritiers qui sans ces alliances demeureraient étrangers les uns aux autres. La parenté de parentèle est constituée à la naissance d’Ego par les unions successives de ses ascendants. La parenté d’affinité, caractéristique des sociétés exogames, résulte de mariages libres avec des étrangers, sans règles ; elle oppose les parents aux affins. La parenté d’alliance, caractéristique des sociétés endogames, implique l’existence de règles, et transforme par l’alliance du mariage les affins en alliés. Bien que différents, ces trois modes convergent sur la conscience d’une ancestralité « traçable », ce qui est un point commun à toutes les sociétés malgaches. L’ancestralité, reconnue comme critère identitaire partout dans l’île, présente une double dimension : généalogique et spatiale, produisant une identité aussi bien individuelle que collective. Ce champ de l’ancestralité structure la société. C’est pourquoi toute fissure dans ce champ déstabilise l’ordre social et peut conduire à des explosions de violence dramatiques.
Plusieurs groupes multidisciplinaires (sciences humaines et sociales, sciences de la vie et médecine) ont engagé un dialogue interdisciplinaire autour des questions sociales et éthiques engendrées par les recherches sur le génome humain (Lee 2008, Caulfield 2009, Takezawa 2014). Afin d’éviter la mécompréhension, la surinterprétation ou la simplification des données générées, sources potentielles de racisme ou d’essentialisme, ces groupes insistent notamment sur deux nécessités : d’une part une attention accrue à accorder à la terminologie désignant les groupes humains étudiés ; d’autre part la responsabilisation des chercheurs qui se doivent d’anticiper les implications sociales et éthiques de la publication et de la diffusion de leurs résultats. Concernant le premier point, l’usage des termes « tribu » ou « ethnie » heurte non seulement le sens moral à cause de leur connotation insidieuse de « race » et des traumatismes historiques auxquels ils renvoient[17][27], mais ce sont aussi des mots imprécis, à géométrie variable. Paul Ottino (1998) en parle ainsi :
il est habituel de répartir les habitants de Madagascar en dix-huit « tribus », un terme qui n’est guère employé que dans ce contexte et dont la transposition en « ethnie » n’est pas plus satisfaisante. Suivant les cas, les appellations supposées désigner des « tribus » ou des « ethnies » correspondent, soit à des milieux écologiques et/ou aux genres de vie particuliers qui s’y rapportent, à d’anciennes configurations politiques ou royaumes (…). En réalité, suivant les critères adoptés, le nombre des ethnies ou tribus malgaches peut varier jusqu’à atteindre la cinquantaine ou bien, à l’inverse, être considérablement réduit. (p. 631)
Sur cette question, plusieurs chercheurs ont tenté de cerner les catégories malgaches qui désignent ces « groupes ethniques », plus particulièrement de préciser les critères de distinction des entités sociales historiques à partir de la désignation vernaculaire de ces dernières[18][28]. Les études voient leurs conclusions converger sur la place centrale d’une origine commune soit sur la base de l’ascendance (ou filiation, firazanana), soit sur la base de l’espace géographique (originaire de, teraky), soit sur la base de l’alliance (matrimoniale ou affinité, firenena). Il y a construction de différences qui s’appuie sur l’ancestralité et sur le territoire, mais qui intègre aussi les opportunités de rapprochement, de contrat avec des groupes ou des individus d’une autre origine ancestrale ou géographique. Les groupes ainsi constitués fonctionnent suivant une hiérarchie sociale verticale dans laquelle la définition statutaire, le rang générationnel et le droit d’aînesse, le genre et la résidence matrimoniale restent des critères importants du pouvoir[19][29]. Dans tous les cas, que ce soit par l’ancestralité, par le territoire ou par les alliances, une parenté est établie ; elle fonde la distinction des groupes. Elle est aussi un des piliers du capital social.
D’autre part, les sociétés malgaches sont toutes hiérarchisées et divisées en ordres sociaux, avec trois groupes statutaires mis en place à la période des royaumes malgaches au 16e siècle : les nobles andriana, les roturiers hova et les esclaves andevo[20][30]. Bien que la colonisation ait aboli l’esclavage, le vécu social ainsi que l’imaginaire collectif ne s’en sont pas détachés, ce qu’expriment plusieurs textes académiques[21][31]. Harisoa Rasamoelina (2012), sociologue, parle de cela en des termes qui, bien que dépassés sur le plan de la structuration sociale[22][32], en traduisent la perception dominante :
à Antananarivo, la bipartition entre mainty et fotsy (Noir/Blanc) est encore actuelle, et chaque groupe s’identifie à partir de traits physiques communs stéréotypés : les descendants d’andevo auraient la peau plus foncée et la chevelure crépue, et les descendants d’andriana et de hova auraient la peau plus claire, et la chevelure lisse. (…) La perpétuation de cette bipartition s’effectue essentiellement dans le cercle familial, avec des consignes bien précises, notamment en ce qui concerne les stratégies matrimoniales. La persistance de cette bipartition et son inscription dans la construction identitaire (…) tient du fait qu’elle a été, et est toujours, un élément essentiel dans les rapports de domination et de pouvoir à Madagascar. (p. 9)
À Madagascar, la terre fait vivre, nourrit, protège, garantit une stabilité sociale, assure la postérité, à un niveau individuel et collectif. Le rapport des hommes à la terre et au territoire sous-tend des rapports sociaux, économiques, religieux et politiques. La terre appartient à l’ancêtre qui le premier l’a défrichée et à ceux qui lui sont affiliés à travers un rapport d’ancestralité ; cette terre ancestrale confère des droits. L’ancestralité transcende à Madagascar la simple généalogie : elle ancre la personne dans un espace, à travers un lien à une terre signifiante, par la voie féminine (cf. teraky[23][33]), et dans un temps, à travers une proximité de l’ascendance, par la voie masculine (cf. firazanana). Or, la généalogie s’appuyant sur l’ADN que propose la génétique est un avatar de cette ancestralité malgache, la matérialisation d’une unité abstraite. Cela appelle d’autant plus à veiller à ne pas décontextualiser l’approche génétique des conditions géographiques (cadre de vie, gestion du sol, etc.) et historiques (gouvernements des milieux, mobilités des hommes, etc.) qui accompagnent la mise en place des territoires et l’installation des sociétés sur lesquelles les études sont conduites. C’est en opérant ainsi que l’approche génétique contribuera à rapprocher toute vérité historique préconçue à la réalité historique des hommes et des lieux où ils vivent. Pour Kimberly TallBear (2008), anthropologue amérindienne,
l’ADN ne doit pas simplement être considéré comme une « chose » moléculaire objective. C’est aussi un dispositif conceptuel à travers lequel les hommes se constituent et déploient des récits organisant la vie : des récits historiques, nationaux, raciaux, des récits sur la famille, la tribu, les origines d’un peuple dont les individus sont eux réels. (p. 236)
Cet ADN que l’homme sensible que nous sommes ne voit pas, ne goûte pas, ne sent pas, est pourtant bien réel lui aussi, mais les récits des généticiens qui le confient à des machines sont des constructions mentales. « ADN, patrimoine génétique », patrimoine matériel ou patrimoine symbolique ? En donnant un peu de leur salive pour en prélever l’ADN, 3000 Malgaches répartis sur l’île tout entière ont récemment accepté de participer à une étude pluridisciplinaire sur le peuplement de Madagascar[24][34]. La salive est un fluide corporel hautement signifiant dans la société malgache : elle symbolise la continuité du flux de vie (aina) (Ottino 1998). Une enquête ethnographique auprès de ces personnes permettrait-elle d’approcher, à travers cette matérialisation qui au fond n’en est pas une, une conception nouvelle de l’ancestralité à Madagascar ? Que leur a-t-on proposé et comment ? Pourquoi ont-elles accepté de participer ? Quels résultats leur a-t-on présentés et comment les ont-elles interprétés ? Quelle conception composite de l’ancestralité produisent-elles ? Quel usage en font-elles? Quelles sont les conséquences de cette étude sur la construction identitaire des personnes et des populations convoquées ?
D’un point de vue éthique, il ne s’agit pas ici d’émettre un jugement de valeur sur la force, la solidité, la souveraineté, la compétence ou la résilience des systèmes de représentations, des croyances, des valeurs morales et des pratiques sociales à Madagascar. Il ne s’agit pas non plus de diaboliser la génétique, mais d’apporter un éclairage sur les recherches en génétique sous le prisme des sciences humaines et sociales, puisqu’il s’agit de génétique « des populations humaines ». Notre posture est d’observer, décrire, essayer de comprendre, partager des interprétations. D’un point de vue épistémologique, ne sommes-nous pas devant le renversement d’un paradigme ? Il ne s’agit plus de la levure, la drosophile ou la souris comme modèle d’étude de l’homme, mais de l’homme comme modèle d’étude de la relation génotype/phénotype, un modèle idéal compte tenu du fait que nous avons accès à son histoire, sa culture, son interaction avec ses environnements écologique, social, culturel… Ce changement de paradigme tend certes à associer à nos comportements des caractères issus de notre patrimoine génétique[25][35], mais ne tend-il pas aussi vers une meilleure considération et documentation de la part non génétique, mais toutefois transmise à nos descendants, de nos apparences et de nos comportements ? Une part héritable par ailleurs incluse dans de nouvelles théories de l’évolution telles que la « synthèse inclusive » (Danchin 2013, Jablonka 2005) ? La morale n’a pas à s’appuyer sur la science. La science n’est pas là pour donner des leçons de vie. Cependant, en alimentant nos réflexions, en induisant un questionnement, en problématisant les situations complexes, l’activité et le discours scientifiques sont opérants, agissants, performatifs ; ils participent de ce monde que nous tentons tous de saisir, les généticiens y compris, ce monde dont la matérialité et l’immatérialité sont indissociables et entretiennent un rapport dialectique complexe.
v
- [Image]: https://www.espacestemps.net/articles/le-nouvel-avatar-ancestralite-dans-historiographie-malgache/figure1-14/
- [1]: #_ftn1
- [2]: #_ftn2
- [3]: #_ftn3
- [4]: #_ftn4
- [5]: #_ftn5
- [Image]: https://www.espacestemps.net/articles/le-nouvel-avatar-ancestralite-dans-historiographie-malgache/figure2-copie/
- [6]: #_ftn6
- [7]: #_ftn7
- [8]: #_ftn8
- [9]: #_ftn9
- Human Genome Project HGP: http://www.genome.gov/10001772
- HUman Genome Organization HUGO: http://www.hugo-international.org
- HapMap: http://www.hapmap.org
- 1000 Genomes: http://www.1000genomes.org
- Human Genome Diversity Project: http://www.hagsc.org
- Genographic Project: https://genographic.nationalgeographic.com
- [10]: #_ftn10
- [11]: #_ftn11
- [Image]: https://www.espacestemps.net/articles/le-nouvel-avatar-ancestralite-dans-historiographie-malgache/figure3-copie/
- [Image]: https://www.espacestemps.net/articles/le-nouvel-avatar-ancestralite-dans-historiographie-malgache/figure4-copie/
- [12]: #_ftn12
- [13]: #_ftn13
- [14]: #_ftn14
- [15]: #_ftn15
- [16]: #_ftn16
- [17]: #_ftn17
- [18]: #_ftn18
- [19]: #_ftn19
- [20]: #_ftn20
- [21]: #_ftn21
- [22]: #_ftn22
- [23]: #_ftn23
- [24]: #_ftn24
- [25]: #_ftn25
